L'origine du génotype des individus
Fiche
Les cellules somatiques de l'individu, obtenues par mitoses à partir de la cellule-œuf, possèdent toutes la même information génétique (aux mutations près). Des cellules de la lignée germinale forment, par méiose, les gamètes qui permettent la reproduction sexuée de l'individu. Comment la reproduction sexuée est-elle à l'origine du génotype des individus ? Comment contribue-t-elle à la diversification du vivant ?
I. La conservation des génomes : stabilité génétique et évolution clonale
La succession de mitoses à partir d'une cellule initiale produit un clone, ensemble de cellules génétiquement identiques entre elles. Ces cellules héritent de l'éventuelle mutation subie par la cellule initiale, et forment alors un sous-clone mutant. Tout accident génétique irréversible est transmis (s'il est viable) par mitoses à la descendance de la cellule. En l'absence d'échanges génétiques avec l'extérieur, la diversité génétique d'un clone résulte de l'accumulation de mutations successives.II. Le brassage des génomes à chaque génération : la reproduction sexuée des eucaryotes
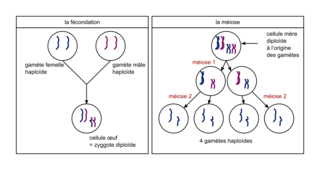
La fécondation est l'union de deux gamètes, mâle et femelle, haploïdes (chacun à n chromosomes, n étant spécifique de l'espèce) et forme une cellule-œuf diploïde, à 2n chromosomes. Les génomes des gamètes ont des origines indépendantes l'une de l'autre, et apportent chacun un lot d'allèles. Dans la cellule-œuf, chaque paire d'allèles est formée de deux allèles identiques (individu homozygote) ou de deux allèles différents (individu hétérozygote).
La méiose permet, à partir d'une cellule initiale diploïde, la formation de 4 gamètes haploïdes. Elle est précédée d'une réplication de l'ADN, au cours de laquelle les chromosomes passent de 1 à 2 chromatides. La méiose est une suite de deux divisions cellulaires : la méiose 1 puis la méiose 2. Lors de la prophase de la méiose 1, les chromosomes homologues d'une même paire s'apparient. En métaphase 1, les chromosomes homologues se disposent de part et d'autre du plan équatorial de la cellule. En anaphase 1, chaque chromosome d'une paire migre de façon aléatoire vers l'un des pôles de la cellule. La télophase 1 aboutit à la formation de deux cellules haploïdes, à n chromosomes à 2 chromatides : chaque cellule contient un chromosome de chaque paire. Puis en méiose 2, dans chaque cellule formée, les chromatides de chaque chromosome se séparent et migrent chacune vers l'une des deux futures cellules filles. Chacun des gamètes est alors haploïde (n chromosomes à 1 chromatide) et contient une copie de chaque gène, soit la moitié de l'information génétique de la cellule initiale. Chez un individu hétérozygote pour un gène étudié, chaque gamète reçoit un seul des deux allèles de ce gène avec une probabilité équivalente, égale à 50 %.
La méiose permet, à partir d'une cellule initiale diploïde, la formation de 4 gamètes haploïdes. Elle est précédée d'une réplication de l'ADN, au cours de laquelle les chromosomes passent de 1 à 2 chromatides. La méiose est une suite de deux divisions cellulaires : la méiose 1 puis la méiose 2. Lors de la prophase de la méiose 1, les chromosomes homologues d'une même paire s'apparient. En métaphase 1, les chromosomes homologues se disposent de part et d'autre du plan équatorial de la cellule. En anaphase 1, chaque chromosome d'une paire migre de façon aléatoire vers l'un des pôles de la cellule. La télophase 1 aboutit à la formation de deux cellules haploïdes, à n chromosomes à 2 chromatides : chaque cellule contient un chromosome de chaque paire. Puis en méiose 2, dans chaque cellule formée, les chromatides de chaque chromosome se séparent et migrent chacune vers l'une des deux futures cellules filles. Chacun des gamètes est alors haploïde (n chromosomes à 1 chromatide) et contient une copie de chaque gène, soit la moitié de l'information génétique de la cellule initiale. Chez un individu hétérozygote pour un gène étudié, chaque gamète reçoit un seul des deux allèles de ce gène avec une probabilité équivalente, égale à 50 %.
La reproduction sexuée : fécondation et méiose
 |
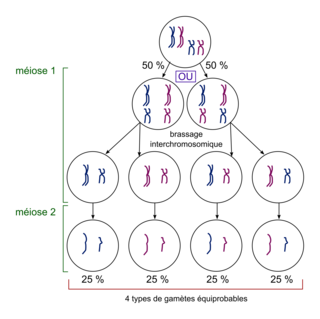
La méiose se caractérise par l'existence de deux brassages génétiques qui augmentent la diversité des gamètes produits. En prophase 1, l'accolement des deux chromosomes d'une même paire permet des crossing-over, des échanges réciproques et équilibrés de portions de chromatides entre deux chromatides appartenant à deux chromosomes homologues. Ce brassage intrachromosomique s'effectue lors de certaines méioses, avec une fréquence variable selon les gènes. En métaphase 1, la disposition de part et d'autre du plan équatorial des chromosomes d'une même paire s'effectue au hasard, conduisant en anaphase 1 à une combinaison aléatoire des chromosomes de chaque paire.
Lors de la méiose, l'étude de deux gènes, présentant chacun deux paires d'allèles, chez un individu hétérozygote montre que les gamètes formés présentent 4 combinaisons d'allèles. Si les deux gènes sont indépendants (situés sur deux chromosomes différents), le brassage interchromosomique aboutit à 4 gamètes équiprobables (fréquence de 25 % chacun). Si les deux gènes sont liés (situés sur le même chromosome), le brassage intrachromosomique forme quatre gamètes non équiprobables. Le nombre de combinaisons génétiques possibles dans les gamètes est d'autant plus élevé que le nombre de gènes à l'état hétérozygote est plus grand chez les parents. La reproduction sexuée, c'est-à-dire la méiose et la fécondation, est à l'origine de la diversité des génotypes des individus d'une même espèce.
Lors de la méiose, l'étude de deux gènes, présentant chacun deux paires d'allèles, chez un individu hétérozygote montre que les gamètes formés présentent 4 combinaisons d'allèles. Si les deux gènes sont indépendants (situés sur deux chromosomes différents), le brassage interchromosomique aboutit à 4 gamètes équiprobables (fréquence de 25 % chacun). Si les deux gènes sont liés (situés sur le même chromosome), le brassage intrachromosomique forme quatre gamètes non équiprobables. Le nombre de combinaisons génétiques possibles dans les gamètes est d'autant plus élevé que le nombre de gènes à l'état hétérozygote est plus grand chez les parents. La reproduction sexuée, c'est-à-dire la méiose et la fécondation, est à l'origine de la diversité des génotypes des individus d'une même espèce.
Les brassages génétiques de la méiose
 |
 |
III. Les principes de base de la génétique
L'analyse génétique est l'étude de la transmission héréditaire des caractères observables, lors de croisements, souvent réalisés entre individus de lignées pures (homozygotes pour tous les caractères) et ne différant que par un nombre limité de caractères. Dans l'espèce humaine, l'identification des allèles portés par un individu repose d'abord sur une étude familiale. Le séquençage de l'ADN permet de connaître le génotype de chaque individu, de ses ascendants et descendants. Les bases de données informatisées permettent d'identifier des associations entre certains gènes mutés et certains phénotypes. Dans les études de type trio, la comparaison de l'ADN d'un sujet porteur d'une mutation avec celui de ses parents permet de savoir si l'enfant a hérité de l'allèle muté de l'un de ses parents ou si la mutation est apparue de novo chez l'enfant.IV. Les accidents génétiques de la méiose
Des anomalies peuvent survenir au cours de la méiose. Lors des crossing-over inégaux, des fragments de chromatides sont échangés de façon déséquilibrée entre chromosomes homologues. Les gamètes formés peuvent ainsi perdre ou gagner certains gènes. Dans certains cas, la duplication de gènes est à l'origine de la formation de familles multigéniques.
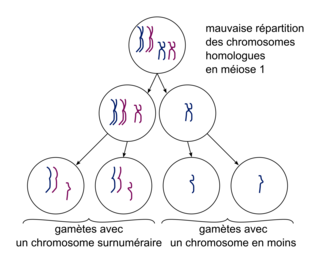
Un autre type d'anomalie est l'absence de séparation des deux chromosomes homologues en anaphase 1, ou des deux chromatides d'un chromosome en anaphase 2, ce qui entraîne l'absence d'un chromosome pour la moitié des gamètes et la présence d'un chromosome surnuméraire pour l'autre moitié. Après fécondation par un gamète normal, les cellules-œufs portent une anomalie chromosomique : soit une monosomie (un chromosome en moins), soit une trisomie (un chromosome en plus). Dans l'espèce humaine, ces anomalies chromosomiques sont fréquemment létales, mais certaines sont viables comme les trisomies 21, 18, et celles affectant les chromosomes sexuels (XXX, XXY ou XYY). D'un point de vue évolutif, les accidents lors de la méiose peuvent être source d'une diversification importante des génomes.
Un autre type d'anomalie est l'absence de séparation des deux chromosomes homologues en anaphase 1, ou des deux chromatides d'un chromosome en anaphase 2, ce qui entraîne l'absence d'un chromosome pour la moitié des gamètes et la présence d'un chromosome surnuméraire pour l'autre moitié. Après fécondation par un gamète normal, les cellules-œufs portent une anomalie chromosomique : soit une monosomie (un chromosome en moins), soit une trisomie (un chromosome en plus). Dans l'espèce humaine, ces anomalies chromosomiques sont fréquemment létales, mais certaines sont viables comme les trisomies 21, 18, et celles affectant les chromosomes sexuels (XXX, XXY ou XYY). D'un point de vue évolutif, les accidents lors de la méiose peuvent être source d'une diversification importante des génomes.
Zoom sur…
Conventions d'écriture en génétique
En génétique, les différents allèles d'un gène sont désignés par une ou deux lettres. L'allèle dominant est écrit en majuscules ou porte le signe (+) en exposant, tandis que l'allèle récessif est écrit en minuscules ou ne porte pas de signe. Par exemple, chez la drosophile, les ailes peuvent être vestigiales (allèle vg) ou normales (allèle vg+). Le phénotype s'écrit entre crochets : une drosophile de phénotype [vg+] possède des ailes normales. Le génotype s'écrit entre parenthèses, et les deux allèles de l'individu sont séparés par deux traits, chacun représentant l'un des deux chromosomes homologues : le génotype d'une drosophile hétérozygote s'écrit (vg+//vg).Croisement-test ou test-cross
Croisement d'un individu présentant le phénotype dominant avec un individu homozygote récessif, c'est-à-dire porteur des allèles récessifs des gènes étudiés. L'étude du phénotype des descendants d'un croisement-test permet de connaître le génotype du parent ayant le phénotype dominant.Dominance et récessivité
Lorsque les deux chromosomes homologues portent des allèles différents pour un même gène, l'allèle qui s'exprime au niveau du phénotype est qualifié d'allèle dominant (par rapport à l'autre allèle). L'allèle qui ne s'exprime pas au niveau du phénotype est dit récessif (par rapport à l'allèle dominant). Lorsque les deux allèles s'expriment conjointement, ils sont dits codominants.Dates-clés
L'histoire de la génétique
1866 : Mendel, moine et botaniste, élabore les premières lois de la transmission des caractères héréditaires.1879 : Fleming observe le comportement des chromosomes au cours de la mitose.
1888 : Strasburger observe le comportement des chromosomes au cours de la méiose.
1901 : De Vries introduit la notion de mutation.
1902 : Sutton et Boveri proposent que les chromosomes soient les porteurs de l'information génétique.
1906 : Johannsen introduit le terme « gène » pour désigner un facteur héréditaire.
1915 : Morgan apporte les preuves expérimentales de la théorie chromosomique de l'hérédité grâce à ses études sur les drosophiles.
1941 : Beadle et Tatum établissent qu'un gène est responsable de la synthèse d'une enzyme.
1944 : Avery, McLeod et McCarty découvrent que l'ADN est le support de l'information génétique chez la bactérie.
1953 : Watson et Crick élaborent le modèle de la structure en double hélice de l'ADN grâce aux travaux de Franklin.
1959 : Lejeune met en évidence l'anomalie chromosomique de la trisomie 21 (syndrome de Down).
1988 : Lancement du projet « Génome humain ».
2003 : Achèvement du séquençage complet du génome humain.
À partir des années 2000 : Mise en évidence de l'importance de l'épigénétique, c'est-à-dire des modifications de l'expression des gènes sans modification de la séquence de l'ADN. Ces modifications sont dépendantes de l'environnement de la cellule, elles sont réversibles et transmissibles à la descendance de la cellule.
© 2000-2026, Miscellane